 ebuhle
commented
4 years ago
ebuhle
commented
4 years ago Hi @kalebentley, thanks for this very informative post. The maps were especially helpful for me.
I'm not in a position to answer your Q1, which as you note depends in part on research questions. But from the modeling perspective, I endorse this suggestion, which would make a hairy problem as tractable as possible:
- Ignore partials and assume all females are fully fecund based on our fecundity by age dataset. This will obviously bias our estimate of egg-to-fry survival low but if the number of partials used is relatively low or the overall proportion of partials is similar among years then the trend in survival shouldn't be impacted too terribly much. At this point, I don't see the purpose of separating North and South channels.
Note that it would still be possible to recover the "true" egg deposition and egg-to-fry survival, if necessary, by post-processing the posterior draws to adjust for the "effective" number of females in each year.
By contrast, option 2 (explicitly modeling partial females) would make the problem even hairier, and would further complicate forecasting because you'd have to specify the effective number of females in advance:
- Designate each partial fish as a relative number of females (e.g., each partial fish counts as 0.75 females). Or perhaps this could be estimated based on our existing dataset? Similar to above, I don't see a point in breaking up the channel if we go this route.
On the data-wrangling front, I think I figured out how to get the raw data -- in general, but specifically for Duncan Channel / Duncan Creek -- into a model-ready format. I'll detail the main points in issue #3, but @kalebentley's post raises a new question / concern:
BYs 2008-10: South and Middle channel re-worked and connected to the top of north channel creating one single channel split by a monitoring weir (south monitoring weir) with another monitoring weir at the bottom (north monitoring weir)
BYs 2011- present: the channel was extended adding more spawning habitat above the south monitoring weir
Do I understand this, and the map, to mean that from outmigration year 2009-present the North weir is counting outmigrants from both South and North channels? So South channel fry are double-counted? For the juvenile data, I'm currently using disposition=="Duncan_Channel" and discarding disposition %in% c("Duncan_North", "Duncan_South"). The former is pretty close to the sum of the latter two, although the equality is not exact in all years. "Duncan_North" is less than "Duncan_South" in some years, which makes me think South channel fry are not double-counted. "Duncan_North" and "Duncan_South" are also missing one year (2014) when "Duncan_Channel" has data. Let me know if "Duncan_Channel" does not represent the total fry output from the channel(s), considered as a single unit.
 kalebentley
kalebentley

 This plot is a bit harder to interpret with all of the groupings but I see a few patterns:
This plot is a bit harder to interpret with all of the groupings but I see a few patterns:




 Hillsont
Hillsont Modified trapping setup at Middle/South channel to deal w/low water.
Modified trapping setup at Middle/South channel to deal w/low water. Combined daily outmigrants at Duncan 2005. Yellow dots are expected emergence dates for fish released into channels based on 1.6K TU accumulation, EF Lewis gauge height for a flow reference.
Combined daily outmigrants at Duncan 2005. Yellow dots are expected emergence dates for fish released into channels based on 1.6K TU accumulation, EF Lewis gauge height for a flow reference.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
Last week, Eric and I had a long phone call to discuss the chum IPM and we spent a fair amount of time talking about the Duncan Creek (i.e., Duncan Springs aka Duncan Spawning Channels) data set. The discussion was spurred by the first round of IPM results where there were clearly some issues and particularly the estimates of freshwater survival seemed unrealistically high. We quickly figured out the Duncan Springs adult data we being "wrangled" incorrectly. Without going into those details, this issue should be easily corrected. However, as I further explained the Duncan dataset and many (but likely not all) of its nuances, Eric and I came to a "crossroads" as to how the Duncan dataset should be evaluated moving forward.
In short, we came up with two general questions for the group:
Below I've tried to provide both some background on the Duncan Channels as well as some very simple summaries of the existing data. NOTE: I wrote this quickly and thus the following text is quite verbose. Read as much or as little as you desire.
For those who are not familiar with the Duncan project, the questions listed above may not make sense so I'll provide a bit of background. In many ways, the Duncan data set is unlike any other chum data set in the Columbia Basin. First and foremost, all adults are manually transported to the spawning area including those that return to the Duncan trap located at the mouth of Duncan Creek. Additionally, the Duncan Spawning Channels are broken into two areas -- north and south (but see notes below) -- which receive female spawners at different reproductive stages. Specifically, each year, one of the two channels is designated to only received "green" females (i.e., females that haven't spawned based on their hard bellies and intact fins) while the other channel can receive "partials" (i.e., females that are either ripe - ready to spawn - or appear to have partially spawn but still believed to have >75% of eggs present based on professional opinion). The channel that receives green females more-or-less alternates each year. The purpose of only placing green females into one of the two channels is so that egg-to-fry survival can be (more) accurately estimated by more accurately estimating the number of eggs deposited in the gravel. By alternating channels, the performance of each "sub-channel" can be monitored and determined if maintenance is needed. In earlier years (BY01 - 05) only green females were used. However, because green females are generally more difficult to "acquire", partials have been used since BY06 to achieve higher spawner abundances in the channels. It should be noted that green females are placed in the partial spawning channel for multiple reasons (see plots below).
As for the Duncan Spawning Channel itself, I'll provide a bit more background that may be useful. Chum salmon were extirpated from Duncan Creek sometime around 1960 when a dam was constructed at the outlet of this creek. In 2000, a fish ladder was installed at the mouth allowing chum salmon access to historical spawning grounds. In September 2001, spawning channels were constructed in the historical spawning area to provide off-channel spawning habitat for chum salmon in Duncan Creek. The spawning channels have gone through several rounds of construction and updates. Todd provided me a long history of the channels and their updates in an email yesterday, which I can share if interested, but here's a brief summary:
For reference, here's an image of the original Duncan Channel configuration (BY2001 - 2007): Here's a Google Earth image from August 2011 of the re-worked channel configuration (BY2008 - 2010). The blue line highlights the old "middle" channel:
Here's a Google Earth image from August 2011 of the re-worked channel configuration (BY2008 - 2010). The blue line highlights the old "middle" channel:
 Here's a Google Earth image from March 2016 of the current Duncan Channel configuration (BY2011 - present):
Here's a Google Earth image from March 2016 of the current Duncan Channel configuration (BY2011 - present):

Using the Google Earth image of the current configuration, the south channel is ~920’ and the north channel is ~200’.
According to Todd, the majority (>80%??) of spawning in the South channel occurs in the most recently constructed portion of the channel (west of the little arm that runs north/south) and very little occurs in the portion of the South (and middle) channel that has existed since 2001. This may partially explain why the productivity of the in the South channel seems so much lower in the
In an effort to better understand the existing datasets, I summarized and generated a couple of plots that may or may not be helpful regarding our questions above.
The first plot shows the number of chum fry outmigrants as a function of female spawners (ignore the specific channels - north or south). The green points signify "green female" years and the blue points signify "partial female" years. Both juvenile (fry) and adult data are essentially censuses in most years. When I squint at the plot, I can potentially see fewer juveniles produced when partial females are used versus green females, which is what I would expect to see

However, when I plotted the number of chum fry outmigrants as a function of female spawners by channel (north vs. south), I no longer see a pattern where the number of outmigrants in "partial years" is lower relative to "green years". This seems counter-intuitive. However, as I mentioned above, green females can be placed in the partial channel and partial females are still supposed to be >75% full of eggs.
Therefore, I was curious about what percentage of females that were placed in each channel were designated as either green (or ripe - eggs coming out but believed to be mostly unspawned) vs. partial. The following plot is similar to the one above except the size of the point is relative to the percentage of spawners that were either green/ripe (i.e., smaller point= more partials). This plot has fewer data points because of the way the chum data are stored - in short, data from BY2012 through 2019 are in a database that is easily queried while earlier years are in spreadsheets. I didn't have time to summarized data from spreadsheets. Nonetheless, what I see is that the north channel has received pretty much fully fecund females each year since BY 2012, even in "partial years" while the south channel (I think by chance) has received a much higher proportion of partials in the partial years. As aside, it also looks like the north channel's capacity could be somewhere around 35K. However, assuming equal spawning distribution and the expected area per redd (2 m2 per female), neither channel should be anywhere close to capacity...
Here's a summary table that includes raw counts of females spawners by brood year (2012 - 2019), channel (north, south), spawner types (green, partial), where the spawners came from, and the reproductive status of each female (G=green, R=Ripe, P=Partial). I'm not sure what's up with the partials being used in the (south) green channel in 2015 and 2017. If needed, Todd and I can look into this later.
Ok - given all of this information, where does this leave us? Here are a couple of options:
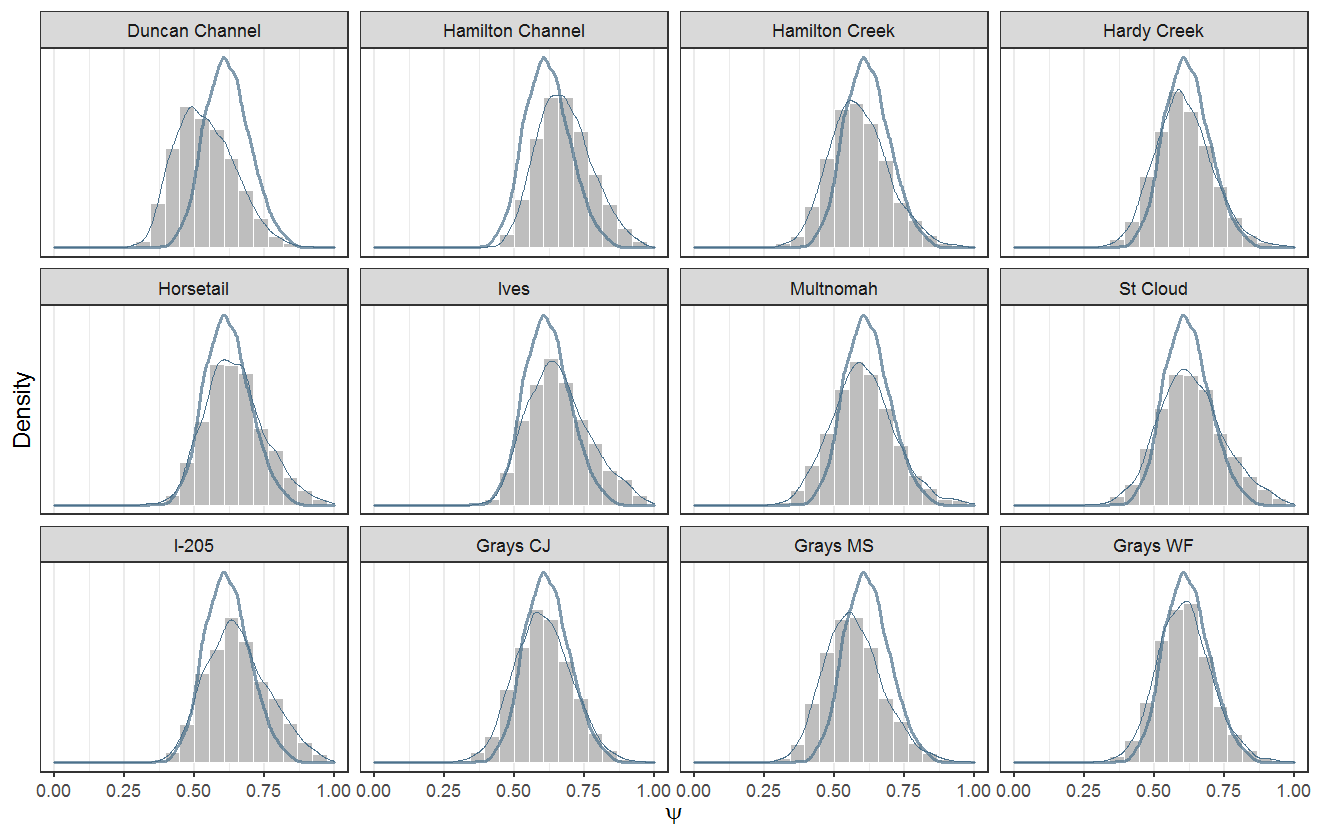
One other thing I haven't done should be sooner-than-later is to compare the estimated egg-to-fry survival rates from the green channel years to the current chum IPM where we are treating Duncan as a single spawning unit and ignoring partials. If the estimates are relatively similar then maybe that tells us that it is unnecessary to sub-divide the monitoring and analysis. However, if there are dissimilar, it would warrant further "analysis" and discussion.